| |
Stachelhäuter, die Bezeichnung "Echinodermata" wurde 1734 durch J.T. Klein eingeführt und benennt einen Stamm der Invertebraten, der ausschliesslich marine Organismen hervorgebracht hat. Echinodermata sind fossil sowie rezent häufig. Als diverse Gruppe haben sie viele ökologische Nischen in verschiedenen Faziesräumen eingenommen. Sie sind im normal salinaren oxischen Milieu weltweit in unterschiedlicher Wassertiefe auf Hart- sowie auf Weichsubstrat verbreitet, wobei die Mehrzahl adulter Echinodermata zum sessilen oder vagilen Benthos des küstennahen Flachmeeres gehört. Finden sie günstige Standortbedingungen vor, treten sie lokal individuenreich in grossen Assoziationen auf. Die Ernährungsweise ist herbivor oder carnivor, selten auch mikrophag. Ihr Verdauungstrakt ist einfach. Das primitive Nervensystem hat kein Gehirn hervorgebracht. Die Vermehrung erfolgt über ein Larvenstadium überwiegend getrenntgeschlechtlich. Echinodermata haben ein Skelett oder Gehäuse aus zahlreichen fest miteinander verbundenen, calcitischen Platten oder aus isolierten Skleriten. Fossile Reste sind sicher seit dem frühen Kambrium aus allen geologischen Systemen bekannt. Als spätproterozoischer Vertreter wird Arkarua, ein Fossil aus Südaustralien, diskutiert. Zu Beginn des Ordoviziums entfalten sich die Echinodermata und durchlaufen eine Blütezeit. Typische paläozoische Formen sind die stieltragenden, sessilen Echinodermata (Pelmatozoen). Früh zeichnet sich ein Evolutionstrend zur Reduzierung des Stiels und der Übergang zur benthisch-vagilen Lebensweise ab, der bis in die Gegenwart erfolgreich ist. Rezente gestielte Echinodermata hingegen haben ihre grosse paläozoische Diversität nicht wieder zurückgewonnen und sich als Reliktformen in grössere Wassertiefen zurückgezogen. Folgende Eigenschaften der Echinodermata sind hervorzuheben: a) Ihr Bauplan beruht wesentlich auf der radialen, pentameren Symmetrie. Die ontogenetisch primäre bilaterale Symmetrie kommt während des bewimperten Larvenstadiums, zurückführbar auf eine einzige Grundform, die hypothetische Dipleurula-Larve, noch zum Ausdruck, wird aber beim adulten Tier überdeckt. b) Echinodermata haben ein hydrostatisches Wasserzirkulations-System (Ambulakral-System, Abb. 1), ein Zusammenschluss von kommunizierenden, flüssigkeitsgefüllten Röhren, auf dem Nahrungsfang, Atmung, Bewegung und sensorische Wahrnehmung wesentlich basieren. Es ist Grundlage für die gerichtete freie Fortbewegung differenzierter Echinodermata (z.B. Seeigel, Seesterne). c) Echinodermata haben ein mesodermal gebildetes Innenskelett bzw. Gehäuse (Calyx, Theka), umschlossen von einer dünnen Haut (Epidermis), das aus unterschiedlichsten, charakteristisch geformten Einzelelementen (z.B. Platten) besteht. Das Wachstum erfolgt durch Ausbildung neuer oder durch Vergrösserung bereits entwickelter Platten, wobei Anwachsstreifen-Muster entstehen können. Beschädigte Skelett-Areale oder abgetrennte Körperanhänge können komplett regeneriert werden. In Abhängigkeit zur Wassertemperatur steht der Einbau von Magnesiumcarbonat in die calcitische Skelettmineralogie. Die Grösse der einzelnen Skelettelemente reicht von weniger als 1 mm Durchmesser (z.B. Holothurien-Sklerite) bis zu mehreren Zentimetern Länge (z.B. Crinoiden-Stielglieder). Unter dem Durchlicht-Mikroskop erscheinen die Einzelplatten oft homogen gelblich oder grau gefärbt und zeigen die typische mikroporöse Maschenstruktur des Stereoms aus stromalen Kanälen, durch die sich zu Lebzeiten das Muskelgewebe zog. Eine eindeutige Bestimmung ist die doppelte Auslöschung als Calcit-Einkristall unter gekreuztem polarisiertem Licht als Folge der Sammelkristallisation, in deren Verlauf sich die kristallographischen C-Achsen der skelettbildenen Kristalle parallel ausgerichtet haben. Postmortal, nach der Zersetzung ihrer Epidermis, zerfallen die Echinodermata-Skelette und werden z.T. gesteinsbildend als isolierte Elemente sedimentiert. Artikulierte Skelett-Komplexe sind fossil selten, oft gebunden an eine rasche Sedimentbedeckung, z.B. durch Tempestite oder Schlammrutschung, und treten eher in dünnbankigen tonigen Kalksteinen und Schiefern (z.B. Posidonienschiefer, Hunsrückschiefer) auf. Weit mehr Volumen nehmen separierte Echinodermata-Reste, insbesondere Pelmatozoen-Stielglieder, vornehmlich in Carbonatgesteinen ein, wo sie eine wichtige Rolle als bioklastische Carbonatproduzenten und Gesteinsbildner spielen, z.B. als Crinoiden-Packstone (Carbonatklassifikation). Dabei können Erhaltungszustand, Sortierungsgrad und Packungsdichte fazielle Hinweise auf Wasserenergie, Sedimentationsrate oder Strömungsrichtung geben. Fossile Echinodermata-Reste gelten als verlässliche Indikatoren für marine Ablagerungsbedingungen. Funktionsmorphologisch sind die Baupläne der Echinodermata stark an ihre Lebensweise und an die Anforderungen ihres Lebensraums angepasst. Sie sind daher besonders für Fragestellungen zur Paläoökologie und Evolution von Bedeutung. Nach einer älteren Klassifizierung wird der Unterstamm Pelmatozoa (gestielte, vorwiegend sessile Echinodermata) den Eleutherozoa (freibewegliche, nicht am Untergrund befestigte Formen) gegenübergestellt. Die neue Klassifizierung basiert auf unterschiedlichen Skelettmerkmalen im adulten Stadium, wobei die taxonomische Stellung vieler Gruppen weiterhin kontrovers ist. Die formenreichsten Gruppen sind folgende:
1) Homalozoa (Carpoidea): ausgestorbene Gruppe mit unsicherer systematischer Stellung, die gewisse Bezüge zu primitiven Chorda-Tieren zeigen. Fossile Funde sind aus dem Mittelkambrium bis Mitteldevon bekannt. Die meisten Gattungen sind aus dem Ordovizium beschrieben. Ihre stark abgeflachte Theka besteht aus calcitischen Plättchen und hat asymmetrische Aussenlinien mit stieloder armähnlichen Fortsätzen. Die meisten Homalozoen lebten als Detritus- oder Suspensionsfresser. Im Gegensatz zu anderen Echinodermata zeigen sie keine Radialsymmetrie und hatten vermutlich auch kein Ambulakral-System.
2) Cystoidea (Beutelstrahler): altertümliche Gruppe, die bereits im Unterkambrium frühe Vertreter stellt und bis ins Karbon nachgewiesen ist. Typisch ist eine gestielte, kugelförmige Theka, von der zwei unverzweigte, armartige Fortsätze (Brachiolen) scheitelwärts abgehen (Abb. 2). Auf der Innenseite der Brachiolen verläuft die Ambulakralfurche, die die Nahrung zum zentral gelegenen Mund leitet. Die Theka besteht aus einem Plattenmosaik, wobei die Platten in Abhängigkeit zur Anordnung und Ausbildung diverse Perforationsmuster von taxonomischem Wert haben.
3) Blastoidea (Knospenstrahler): Im Gegensatz zu den Crinoiden, mit denen sie während des Paläozoikums eine ökologische Nische als Suspensionsfiltrierer, meist in gemässigt durchströmten Habitaten, teilten, sterben sie an der Wende zum Mesozoikum aus. Erfolgreich waren sie v.a. in Riffstrukturen des Unterkarbons. Das Gehäuse besteht aus einer knospenförmigen Krone (Calyx), (Abb. 3), die meist einem Stiel aufsitzt, mit dem die Blastoideen fest auf dem Untergrund aufgewachsen sind. Die verhältnismässig kleine Calyx besteht bei differenzierten Formen aus 13 Platten, welche radialsymmetrisch in drei Reihen angeordnet sind. Ihr Mund liegt gegenüber der Stiel-Anwachsstelle am Top der Krone, der After lateral. Von den Ambulakra erstrecken sich die dünnen unverzweigten Brachiolen, die aus biserial angeordneten Platten bestehen. Vermutlich fingen sie ihre Nahrung mit Schleimfilmen oder Ciliae, ohne Ambulakral-Füsschen. Eine Perforation, wie die der Cystoideen-Platten, fehlt.
4) Crinoidea (Seelilien): seit dem Ordovizium belegte Gruppe von Suspensionsfiltrierern, die besonders auf paläozoischen Carbonatplattformen weit verbreitet war. Rezent sind noch mehrere 100 Arten aus dem Subtidal der tropischen bis polaren Regionen sowie der Tiefsee bekannt. Adult leben sie meist festgewachsen auf Substrat, seltener auch pseudoplanktonisch oder planktonisch-pelagisch (seit Oberjura). Stiellose Flachwasser-Formen können sich mit Cirren-Fortsätzen anheften. Typische Tiefwasserformen sind gestielt. Generell besteht der Körper aus einem gegliederten Stiel (Columna) und einer Krone (Kelch mit fünf oder mehr Armen, Abb. 4). Der Stiel kann sekundär zurückgebildet sein oder an seiner Basis zusätzliche Verankerungen (Stielwurzeln) ausgebildet haben. Die einzelnen Stielglieder haben runde, ovale oder sternförmige Umrisse und spezielle Muster an den Gelenkflächen. Stielglieder überwiegen fossil quantitativ. Die Weichteile werden von einem Kelch aus Dorsalkapsel und Kelchdecke (Tegmen) umschlossen. Die Calyx setzt sich aus zwei oder drei Plattenreihen zusammen, wobei sich an der oberen Reihe die gelenkartigen Ansatzstellen der Arme befinden. Diese, meist federartig verzweigt, erzeugen einen Nahrungspartikelstrom, der über die auf den Innenseiten der Arme verlaufenden Ambulakral-Rinnen mit Hilfe der Ambulakral-Füsschen direkt abwärt zum Mund am Top der Calyx geleitet wird.
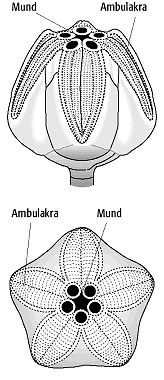
5) Edrioasteroidea: Sie treten vom Unterkambrium bis ins Karbon auf und stellen eine primitive Gruppe der Echinodermata dar. Als präkambrische Vorläuferform wurde die triradialsymmetrische Form Tribrachidium der Ediacara-Fauna in Betracht gezogen. Edrioasteroideen-Skelette bestehen aus einer scheibenförmigen Theka (6-60 mm im Durchmesser) mit zentral gelegenem Mund. Von diesem ziehen sich fünf Nahrungsrinnen (Ambulakra) meist sternförmig nach aussen, die jeweils durch Deckplatten geschützt sind (Abb. 5). Brachiolen sind nicht ausgebildet.
6) Echinoidea (Seeigel) : seit dem Ordovizium bekannte Gruppe, die benthisch vagil, küstennah bis in Tiefsee-Bereiche, auf Weich- oder Hartsubstrat, grabend oder auf der Sedimentoberfläche lebt. Ihr Optimum finden sie im subtropischen bis tropischen Litoral, wo sie besonders artenreich auftreten. Ihr starres Gehäuse (Corona) besteht aus zahlreichen miteinander verbundenen calcitischen Tafeln, die mit beweglichen Stacheln oder Pedicellarien (zangenartige Greifwerkzeuge) ausgestattet sein können, welche dem Nahrungsfang, der Fortbewegung und der Abwehr dienen. Arme und Stiel fehlen. Sie besitzen ein differenziertes Ambulakral-System, ähnlich dem der Seesterne. Zwei Organisationsformen werden unterschieden: a) Bei den regulären Seeigeln liegen sich After und Mund im vertikalen Gehäusequerschnitt gegenüber (Abb. 6). Ihr radialsymmetrisches Gehäuse ist meist hemisphärisch geformt und ihre Stacheln sind gleichmässig auf der Oberfläche verteilt. Der After liegt auf der dorsalen Gehäuseseite, im Periprokt, einem membranartigen Gebiet im Zentrum des Apikalfeldes. Diese Scheitelregion besteht meist aus jeweils fünf Ocellar- und fünf Genital-Platten (einschliesslich Madreporen-Platte), die rosettenartig angeordnet sind. An das Apikalfeld schliessen sich vertikal im Wechsel jeweils fünf Reihen Interambulakral- und Ambulakral-Felder radialstrahlig an. Diese setzen sich aus paarig angeordneten, alternierend geformten Platten zusammmen. Aus den perforierten Ambulakral-Platten ragen die Ambulakral-Füsschen. Auf der ventralen Gehäuse-Seite sitzt das Peristom, welches meist aus einer lederartigen Membran besteht, in dessen Zentrum der Mund liegt. Es beinhaltet den Kauapparat, die sogenannte "Laterne des Aristoteles", der aus 40 Einzelelementen besteht und eine Ernährungweise als Oberflächen-Weider oder Aasfresser unterstützt. Fossil sind reguläre Echinoideen häufig durch einzelne Skelettelemente, als Bioklasten, überliefert. Im Dünnschliff zeigen die Stacheln typische Umrisse und Querschnitte mit radialstrahligen Mustern. b) Im Zuge der adaptiven Radiation im Unterjura treten die irregulären Seeigel hinzu, bei denen es zur Verlagerung des Mundes nach vorne und des Afters in eine hintere Position des Interambulakralfeldes, ausserhalb des Apikalfeldes, kam. Typisch ist ein bilaterales, abgeflachtes Gehäuse mit kleineren Stacheln. Das Peristom-Gebiet ist reduziert und der Mund durch eine Lippe geschützt. Viele leben grabend und tragen dadurch in erheblichem Umfang zur Bioturbation bei. Dabei bewegen die Organismen das Sediment vornehmlich mit den vorderen Stacheln von den Flanken zur Seite. Einige herzförmige Formen (Spatangoideen) können sich bis zu 20 cm tief in das Substrat eingraben. Dabei wird ein senkrechter Kanal von Sediment freigehalten, der die Zufuhr von Atemwasser gewährleistet. Aufgrund dieser Lebensweise werden sie fossil oft in situ gefunden. Hervorzuheben ist die Bedeutung der Echinoideen für die Zonengliederung in der Kreide.
7) Holothuroidea (Seegurken, Seewalzen): Sie sind seit dem Ordovizium bekannt. Rezent existieren über 1000 Arten. Sie besiedeln ein weites Spektrum an Lebensräumen, wo sie vagil-benthisch, in Ausnahmen pelagisch vorkommen. Als einzige Gruppe der Echinodermata sind sie euryhalin und haben auch den brackischen Lebensraum erschlossen. Ihr zylindrischer, muskulöser Weichkörper hat keine Arme. Um den Mund befindet sich ein Tentakelring, dessen Umriss bedeutsam für die Rezent-Taxonomie ist. Ambulakral-Rinnen verlaufen längs über den sekundär bilateral-symmetrischen Körper. Extreme Flexibilität und Beweglichkeit des Körpers ist die Folge der Reduzierung des artikulierten Skeletts auf winzige ( < 1 mm), fragile Sklerite, die in einer lederartigen Haut sitzen. Vollständige Fossilien sind äusserst selten. Weit verbreitet sind Funde einzelner Holothurien-Sklerite, die grossen Formen- und Ornamentreichtum besitzen (z.B. Haken, Anker, Speichenrädchen) und präparativ durch Säureaufbereitung als Mikrofossilien aus Carbonaten gewonnen werden. In der Alpinen Trias haben die Sklerite biostratigraphischen Wert.
8) Asteroidea (Seesterne): Sie sind seit dem Ordovizium nachgewiesen und haben einen sternförmigen Körperumriss von einigen Millimetern bis 50 cm im Durchmesser mit fünf oder mehr Armen. Auf der flachen Unterseite der Arme verlaufen die Nahrungs-Rinnen mit den Ambulakral-Füsschen radialstrahlig zum zentralen Mund. Der After liegt auf der Körperoberseite. Ein differenziertes Wasserzirkulations-System ermöglicht eine aktive, freie Bewegung. Im einzelnen erfolgt die Anpassung des Wasservolumens über den Steinkanal, an dessen Ende eine Einzelöffnung (Hydropore) oder eine siebartige Genitalplatte (Madreporenplatte) die Flüssigkeitsabgabe bzw. -aufnahme steuert. Vom Ringkanal, welcher mit dem Steinkanal in Verbindung steht, gehen fünf Radialkanäle ab, die wiederum verbunden sind mit kurzen lateralen Seitenzweigen, die zu den Ampullen und schliesslich zu den Ambulakralfüsschen führen. Radialkanäle, laterale Fortsetzungen, Ampullen und Ambulakralfüsschen sind Elemente des Ambulakrum. Manche Seesterne haben eine räuberische Lebensweise entwickelt. Zur Beute gehören u.a. Muscheln, deren Klappen sie durch das Ziehen der festgesaugten Ambulakral-Füsschen zum Klaffen bringen. Beisswerkzeuge sind nicht entwickelt, sie können ihren Magen ausstülpen und verdauen die Beute extern. Lokales Massenauftreten der rezenten Gattung Acanthaster (Gruppe der Dornenkronen) stellt heute infolge der anthropogenen Dezimierung ihres Fressfeindes eine Bedrohung der Riffkorallen im Indopazifik dar.
9) Ophiuroidea (Schlangensterne): Seit dem Unterordovizium bekannte vagil-benthische Gruppe. Im Gegensatz zu den Seesternen besitzen sie deutlich abgesetzte Arme, die sich von der zentralen Körperscheibe aus radialstrahlig erstrecken. Die zahlreichen Wirbel der Arme sind gelenkartig miteinander verbundenen und gewährleisten eine hohe Beweglichkeit. Der sternförmig geschlitzte Mund liegt auf der Körperunterseite (oral) und trägt fünf Kiefernpaare, die von den Buccalschildern gesäumt sind. Der Mund übernimmt die Funktionen des nicht separat ausgebildeten Afters. Die Madreporenplatte liegt meist aboral.
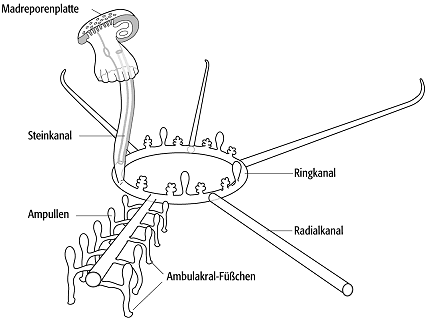 Echinodermata 1: Ambulakral-System rezenter Seesterne mit Madreporen-Platte, Steinkanal, Ringkanal und Radialkanälen. Ambulakral-Füsschen und Ampullen sind nur für einen Arm gezeichnet. Echinodermata 1: Ambulakral-System rezenter Seesterne mit Madreporen-Platte, Steinkanal, Ringkanal und Radialkanälen. Ambulakral-Füsschen und Ampullen sind nur für einen Arm gezeichnet.
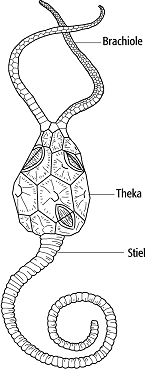 Echinodermata 2: Cystoidea, Pleurocystites, rundliche Theka mit Stiel und paarigen, unverzweigten Brachiolen. Echinodermata 2: Cystoidea, Pleurocystites, rundliche Theka mit Stiel und paarigen, unverzweigten Brachiolen.
 Echinodermata 3: Blastoidea, Pentremites, Karbon, knospenförmige Theka (ca. 3 cm lang) mit langen Ambulakra und zentralem Mund. Echinodermata 3: Blastoidea, Pentremites, Karbon, knospenförmige Theka (ca. 3 cm lang) mit langen Ambulakra und zentralem Mund.
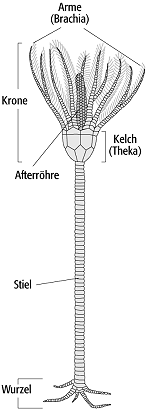 Echinodermata 4: Crinoidea, dreigeteilter Grundbauplan, Wurzel, Stiel und Krone. Die Krone besteht aus Kelch und Armen. Echinodermata 4: Crinoidea, dreigeteilter Grundbauplan, Wurzel, Stiel und Krone. Die Krone besteht aus Kelch und Armen.
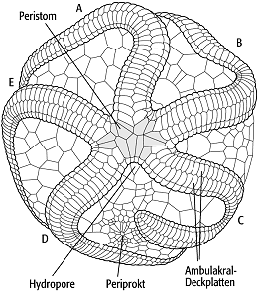 Echinodermata 5: Edrioasteroidea, Edrioaster, Ordovizium, Oberflächenansicht mit Ambulakra unter Deckplatten, Periprokt mit Mund und Peristom. Echinodermata 5: Edrioasteroidea, Edrioaster, Ordovizium, Oberflächenansicht mit Ambulakra unter Deckplatten, Periprokt mit Mund und Peristom.
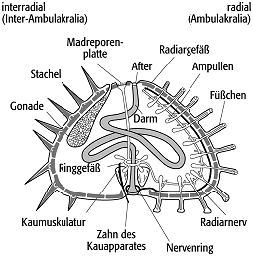 Echinodermata 6: Echinoidea, Grundbauplan eines regulären Seeigels mit gegenüberliegender Position von After und Mund. Echinodermata 6: Echinoidea, Grundbauplan eines regulären Seeigels mit gegenüberliegender Position von After und Mund. |
|
