| |
[von lat. evolvere=ab-, entwickeln], 1) umgangssprachlich: Entwicklung, Umwandlung, auch Höherentwicklung; 2) im engeren Sinne: die Biosphäre verändert sich ständig, weil das Erbgut in einem gewissem Masse instabil ist. Bei der Weitergabe der Erbinformationen, die in den Genen lokalisiert und durch die DNS codiert sind, schleichen sich Kopierfehler ein, die einen permanenten Umbau des Genoms (Mutationen) und der Steuermechanismen ermöglichen und so zu einem ständigen Wandel führen. Damit ist die Voraussetzung für eine Evolution gegeben. Der Umbau des Genoms führt nicht zu einem Chaos, weil strenge Kontrollen den Veränderungen enge Grenzen setzen. Schon bei der Rekombination der Chromosomen in der Eizelle wird die Entwicklung des Keims bei grossen Veränderungen im Genom abgebrochen. Ein Individuum kann auch später nur überleben, wenn es in jeder Phase lebensfähig ist, d.h. es wird in jedem Stadium einer strengen Funktionskontrolle durch die Umwelt unterworfen. Die genetischen Veränderungen sind zunächst ungerichtet und auf dem molekularen Niveau auch reversibel. Bislang sind keine inneren Vorgänge bekannt, die der Evolution eine Richtung geben. Von aussen wirken allerdings Selektionsmechanismen, die aus der Variabilität zwischen den Individuen einer Population diejenigen bevorzugen, die in dem entsprechenden System besser angepasst sind. Das bedeutet, dass sie über ein besseres Ressourcen/Leistungsverhältnis verfügen. Dadurch wird eine Richtung vorgegeben, in der die Leistungssteigerung als Selektionsvorteil eine wesentliche Komponente bildet. Eine Umkehr grösserer Evolutionsschritte würde eine Verringerung der Leistung bedeuten und ist damit unwahrscheinlich. Da nicht alle Leistungssteigerungen in der gleichen Richtung verlaufen müssen, wird eine stärkere Differenzierung der Lebensformen möglich, die sich in einer immer grösseren Diversität äussert.
Evolutionsvorgänge wurden seit dem Beginn des 19. Jh. vielfach diskutiert, doch erst in der Evolutionstheorie, die Charles Darwin 1859 formulierte, wurde der Mechanismus für die Evolution im Zusammenspiel von angebotener Vielfalt und Selektion erkannt. Seit Darwin sind viele Ergänzungen und Detailaspekte hinzugekommen, aber seine Grundidee bildet noch immer die beste Möglichkeit, die umfangreichen Befunde aus Biologie und Paläontologie sinnvoll zu erklären. Allerdings kann und will diese Evolutionstheorie keine Antworten auf die Sinnfragen geben, wie sie von Religion und Philosophie gestellt werden, weil die Naturwissenschaften den Menschen zwar in ihr Weltbild einschliessen, ihn aber nicht zum Ausgangspunkt der Betrachtung machen und ihm nicht die zentrale Stellung einräumen. Die Evolution der einzelnen Tier- und Pflanzengruppen ist an einen konkreten zeitlichen und räumlichen Rahmen gebunden und damit ein historischer Vorgang, der nur anhand von Fossilien überprüft werden kann.
Schon die Tatsache, dass die einzelnen Wirbeltierklassen in der Erdgeschichte nacheinander auftreten, bietet einen unübersehbaren Hinweis auf die Evolution. Die meisten Tierstämme haben erst im Kambrium mineralisierte Hartteile entwickelt und sind von diesem Zeitpunkt an im Fossilbericht erkennbar. Ihre frühe Evolution muss daher weitgehend nach der Methode der vergleichenden Anatomie erschlossen werden. Die Wirbeltiere erscheinen erst im Paläozoikum. Der Fossilbericht zeigt sehr eindrucksvoll das Auftreten immer höher differenzierter Klassen in einer zeitlichen Abfolge (Abb.). Im Ordovizium treten die Fische auf, zuerst kieferlos, später kiefertragend. Im Oberdevon lassen sich die ersten Amphibien und im Karbon die ersten Reptilien nachweisen. Aus der Obertrias sind die ersten Säugetiere bekannt, während die Vögel erst seit dem Oberjura belegt sind. Diese Höherentwicklung kann jeweils mit der Eroberung neuer Lebensräume gleichgesetzt werden. So gelingt den Amphibien der Schritt auf das Land. Die Reptilien sind auch bei der Eiablage nicht mehr an das Wasser gebunden. Die Säugetiere können als warmblütige Tiere auch nachts aktiv sein, und die Vögel erobern den Luftraum. Der Begriff "Höherentwicklung" muss stark relativiert werden, weil Vögel und Säugetiere sich aus unterschiedlichen Reptilgruppen in verschiedene Richtungen entwickelt haben. Daher kann sich ein sinnvoller Vergleich der Entwicklungshöhe stets nur auf einzelne Organe, nicht aber auf den ganzen Organismus beziehen.
Das zeitliche Auftreten der einzelnen Wirbeltierklassen ist nur ein gewisser Hinweis auf die Evolution, überzeugender ist der Nachweis von Übergangsformen in der entsprechenden stratigraphischen Position. Die berühmteste Zwischenform ist der Urvogel Archaeopteryx aus der Lagune von Solnhofen. Dieses Tier besitzt mit der Bezahnung und dem langen Schwanz typische Reptilmerkmale, während die Flugfedern typisch für Vögel sind. Archaeopteryx bietet mit diesem Merkmalsmosaik ein gutes Modell für einen Übergang. Allerdings ist Vorsicht geboten, in Archaeopteryx den Stammvater aller Vögel zu sehen, weil der Fossilbericht recht lückenhaft ist. Es ist durchaus denkbar, dass die Vögel etwas früher entstanden sind, und im Oberjura an anderer Stelle bereits höher entwickelte Formen existierten. Dennoch bleibt Archaeopteryx das beste Modell für den Übergang von den Reptilien, speziell den theropoden Dinosauriern, zu den Vögeln, bis weitere Funde die bestehenden Lücken verringern.
Die Evolution lässt sich im Fossilbericht noch eindrücklicher auf dem Gattungs- bzw. Artniveau verfolgen. Mikrofossilien oder Kleinsäuger sind besonders geeignet, weil sie in grossen Zahlen vorliegen. Betrachtet man Populationen aus einer zeitlichen Abfolge, dann lässt sich eine Merkmalsverschiebung statistisch verdeutlichen. So verändern die Foraminiferen der Oberkreide Norddeutschlands ihre Breite und die Höhe kontinuierlich. Gelegentlich wird dabei auch die Aufspaltung in verschiedene Taxa deutlich. Bei den ursprünglichen Primaten, die im Alttertiär von Nordamerika sehr häufig sind, kann man eine systematische Verschiebung der Merkmalsvariabilität am schrittweisen Umbau des Gebisses ablesen. Daraus ergibt sich sowohl die Abgrenzung aufeinanderfolgender Arten, wie die Aufspaltung in verschiedene Entwicklungslinien. Ähnliche Umkonstruktionen werden überall sichtbar, sobald die Fossilien von Tier- oder Pflanzengruppen aus einem längeren Zeitraum miteinander verglichen werden.
In den letzten Jahren wurde intensiv diskutiert, ob die Evolution sprunghaft grössere Veränderungen hervorbringt (Punktualismus) oder ob sich die Formen gleichmässig verändert haben (Gradualismus). Der Fossilbericht zeigt meist einen gleichmässigen Umbau. Aus der Gesamtsicht lassen sich jedoch grosse Unterschiede in der Entwicklungsgeschwindigkeit zwischen den Taxa postulieren. "Lebende Fossilien", wie der Pfeilschwanzkrebs Limulus, sind durch eine sehr geringe Veränderungsrate charakterisiert. Obwohl die Evolutionsgeschwindigkeit schwer zu quantifizieren ist, kann sie selbst innerhalb einer Evolutionslinie sehr erheblich wechseln. Fledermäuse tauchen im Eozän nahezu fertig evoluiert auf, wie die vorzüglichen Funde aus der Grube Messel bei Darmstadt zeigen. Ihre Evolution in diese spezielle Nische hinein dürfte relativ schnell erfolgt sein, danach aber waren die Möglichkeiten für eine leistungssteigernde Umkonstruktion sehr begrenzt. Deswegen zeigen sie in den letzten 45 Mio. Jahren fast keine weiteren Veränderungen mehr. Andere Säugetiergruppen entwickeln sich erst in diesem Zeitraum und zeigen ganz andere Phasen gesteigerter Entwicklung. Die Primaten beginnen z.B. relativ langsam und weisen erst in der jüngsten Erdgeschichte eine höhere Evolutionsgeschwindigkeit auf. Die Evolution scheint sich als historischer Vorgang aller Schematisierung zu entziehen. Die Biostratigraphie benutzt den sich ständig wechselnden Artbestand in Fauna und Flora, um aus dem Vergleich relative Altersangaben zu gewinnen. Besonders hilfreich sind dabei jene Arten, die sich sehr schnell entwickelten und gleichzeitig eine sehr weite Verbreitung hatten (Leitfossil).
Für die Rekonstruktion der Phylogenie (Stammesgeschichte) einzelner Gruppen gibt es sehr unterschiedliche Verfahren. Der stratigraphische Ansatz erscheint zunächst logisch. Wenn in einer Gegend eine ältere und eine jüngere Form der gleichen Gruppe vorliegen, dann sollte die jüngere höher evoluiert sein und von der älteren abstammen. Dieser Ansatz ist aber nur begrenzt brauchbar, weil der lokale Aspekt zu sehr im Vordergrund steht und mögliche Arealverschiebungen unberücksichtigt lässt. Da Entwicklungsfortschritte oft in kleineren Randpopulationen zu erwarten sind, muss man auch damit rechnen, dass eine evoluiertere Form einwandert, ohne dass sie von der primitiveren Form der betrachteten Region abstammt. Bei den tertiären und pleistozänen Pferdeverwandten aus Europa zeigen die Gattungen Anchitherium, Hipparion und Equus jeweils ein höheres Evolutionsniveau. Dennoch bilden sie keine echte Evolutionsreihe, weil jede dieser Gattungen zu anderen Zeiten aus Nordamerika eingewandert ist. Die eigentliche Evolution der Pferde fand nämlich in Nordamerika statt, von wo sich mehrfach Formen über Asien nach Europa ausgebreitet haben. Grundsätzlich lässt sich aus der Ähnlichkeit zwischen Taxa der Verwandtschaftsgrad ablesen. Die vergleichende Anatomie hat seit dem frühen 19. Jh. mit diesem Ansatz ganz grundlegende Erkenntnisse über die Verwandtschaftsverhältnisse geliefert. Bis etwa 1960 betrachtete man vorwiegend die allgemeine Ähnlichkeit, dabei blieb es der Erfahrung überlassen, welchen Merkmalen ein besonderes Gewicht beigemessen wurde. Wesentliche systematische Fortschritte wurden dadurch erzielt, dass man streng zwischen gemeinsamen Merkmalen, die primitiv (plesiomorph) sind und deswegen in einer grösseren Verwandtschaftsgruppe auftreten, und abgeleiteten (apomorphen) Merkmalen, die nur ein Taxon kennzeichnen, unterschied. Die Merkmale, die eine eng verwandte (monophyletische) Gruppe von anderen Formen abgrenzen, sind synapomorph. Nur die gemeinsamen abgeleiteten (synapomorphen) Merkmale können eine engere Verwandtschaft belegen. Dabei ist allerdings die Unterscheidung von plesiomorphen und apomorphen Merkmalen keineswegs einfach, manchmal sogar etwas subjektiv, und hängt immer von den betrachteten Arten ab. Etwa seit 1990 kann man nicht nur morphologische Strukturen, sondern auch die molekularen Sequenzen aus der DNS vergleichen. Es lassen sich damit auch weiter voneinander entfernt stehende Gruppen vergleichen, zwischen denen ein morphologischer Vergleich nicht mehr sinnvoll ist. Zwar kann man bei der molekularen Konfiguration nur selten zwischen primitivem und abgeleitetem Zustand unterscheiden, aber wegen der grossen Zahl der Daten ist es möglich, einen hypothetischen Entwicklungsweg zu berechnen, der die wenigsten Änderungen erfordert (Sparsamkeitsprinzip). Dabei werden die Regeln des Kladismus angewendet. Die sich ergebenden Stammbäume müssen jedoch stets sorgfältig geprüft werden, weil der sparsamste Weg zwar logisch zu ermitteln ist, aber nicht unbedingt dem historischen Weg zu entsprechen braucht. Alle diese Methoden setzen voraus, dass Konvergenzen, das sind die Ähnlichkeiten, die nicht auf Verwandtschaft beruhen, unberücksichtigt bleiben. Konvergenzen sind besonders dann auffallend, wenn einfache biomechanische Gegebenheiten zur Selektion ähnlicher Formen geführt haben. Eine hydrodynamisch günstige Körperform ist mehrfach entwickelt worden, wenn ein schnelles, dauerhaftes Schwimmen vorteilhaft war. Bei komplizierteren Strukturen, wie etwa dem Flügel, den Flugsaurier, Vögel und Fledermäuse unabhängig voneinander aus der Vorderextremität entwickelt haben, lässt sich der unterschiedliche Ursprung an den abweichenden Konstruktionen erkennen. Die Evolution ist ein historischer Vorgang, d.h. eine Veränderung in Zeit und Raum. Die einzelnen Schritte in der Evolution von Pflanzen und Tieren erfolgen nicht isoliert, sondern sind in hohem Masse vom biologischen Umfeld abhängig. Zum einen bedingen sich Taxa gegenseitig, wie Blütenpflanzen und Insekten (Co-Evolution), zum anderen gibt es einmalige Evolutionssituationen. Dazu gehört die Eröffnung eines neuen Lebensraumes. Als die Quastenflosser das Land erstmals besiedelten, gab es dort noch keine Konkurrenten. Bei jedem späteren Versuch das Land erneut zu besiedeln, warteten dort bereits räuberische Tiere. Auch geographische Voraussetzungen führen zur Einmaligkeit der Evolution. Die Abgeschlossenheit von Inseln oder isolierten Kontinenten schafft für Landtiere besondere Verhältnisse. In Madagaskar und Südamerika haben sich über lange Zeit stark endemische Formen entwickelt. Deswegen ist die Evolution zwar ein überall wirkender Prozess, der sich aber in seinen Ergebnissen sehr stark in Raum und Zeit differenziert. Für dieses historische Geschehen bildet der Fossilbericht die wichtigste Quelle. WvK
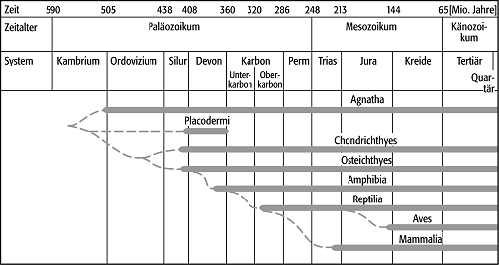 Evolution: Die Klassen der Wirbeltiere treten nacheinander im Fossilbericht auf und lassen eine Entwicklung zu komplexeren Strukturen erkennen. Evolution: Die Klassen der Wirbeltiere treten nacheinander im Fossilbericht auf und lassen eine Entwicklung zu komplexeren Strukturen erkennen. |
|
