| |
ausgestorbene Tiergruppe, die vom Kambrium bis ins obere Perm vertreten war. Ihre Blüte erreichte sie im Oberkambrium und Ordovizium. Die meisten der ausschliesslich marinen Organismen lebten benthisch (benthische Organismen), wobei sie hauptsächlich in kalkigen Plattformsedimenten oder feinkörnigem Detritus anzutreffen sind. Manche Formen kommen aber auch in tieferem Wasser, z.B. an Kontinentalhängen bzw. in pelagischen Zonen vor. Die Körpergrösse schwankt i.a. zwischen 3 und 10 cm, es gibt aber auch bis ca. 70 cm grosse Formen sowie Arten, die <5 mm sind und damit zu den Mikrofossilien gehören.
Trilobiten schützten ihren Körper durch einen Panzer, dessen auffällige Dreiteilung namengebend für die ganze Gruppe war (Dreilappkrebse) (Abb. 1). In der Längsrichtung unterscheidet man die Rhachis als medianen Bereich sowie die beiden lateralen Zonen, die Pleuralbereiche. Quer gliedert sich der Panzer in Cephalon, Thorax und Pygidium. Seitlich ist der Panzer nach unten umgeschlagen und bildet dort die sog. Duplikatur, die aber nicht der Duplikatur der Ostracoda entspricht. An ihr setzt der Weichkörper an. Das in etwa halbkreisförmige Cephalon ist ein weitgehend ungegliederter Schild, der aus der mehr oder weniger vollständigen Verschmelzung der Kopfsegmente resultiert. Bei manchen Gruppen kann der Kopfschild enorm vergrössert sein. Ist dieser Bereich perforiert, bezeichnet man ihn als Siebhaube. Als Fortsetzung der Rhachis bildet die Glabella eine zentrale Aufwölbung. Ausdruck der Weichkörpersegmentierung ist die Glabellarfurchung, die jedoch auch rückgebildet sein kann. Die unterste Furche ist allerdings stets durchlaufend und bildet den sog. Occipital- oder Nackenring. Die Bereiche rechts und links der Glabella heissen Wangen. Häutungsnähte teilen die Wangen in Fest- und Freiwangen. Die Festwangen schliessen unmittelbar an die Glabella an; sie bilden mit ihr das Cranidium, während die Freiwangen inklusive möglicher Wangenstacheln durch den Häutungsprozess vom Cranidium getrennt werden (Abb. 2). Auch die Augen selbst gehören meist zum Bereich der Freiwangen. Bei den Lichtsinnesorganen unterscheidet man das holochroale und das schizochroale Auge. Der holochroale Typus ist das primitivere, aber am weitesten verbreitete Organ. Hier werden bis zu 15.000 Linsen von einer gemeinsamen Cornea (Hornhaut) überdeckt, während das schizochroale Auge über weniger, aber dafür Einzellinsen verfügt. Auf der Unterseite des Kopfes befinden sich das vor oder über der Mundöffnung liegende Hypostom und das sich manchmal nach unten anschliessende Metastom als weitere Panzerteile. Das Hypostom kann taxonomische Bedeutung haben. Vor dem Hypostom liegt die spangenförmige Rostralplatte; sie ist manchmal mit dem Hypostom verwachsen. Der Thorax setzt sich aus einer Reihe beweglicher Segmente zusammen, die zwischen 2 und 44 liegt. Zumindest auf Familienniveau ist die Anzahl der Segmente meist konstant. Jedes Thorakalsegment ist aus dem medianen Axialring sowie den beiden seitlichen Pleuren aufgebaut. Die Pleuren haben oft gelenkige Verbindungen. Mit Hilfe von Fortsätzen, die in Gruben vorhergehender Pleuren passen, werden Verschiebungen verhindert. Jeder Axialring trägt an seinem vorderen Rand einen sog. gelenkigen Halbring, der nur bei der Einrollung zu sehen ist. Er deckt die durch Körperdehnung bedingte freiliegende Stelle ab. Der hintere Körperabschnitt ist das Pygidium und besteht aus einer einheitlichen Platte. Das ursprünglich sehr kleine Pygidium wurde durch Angliederung von Thorakalsegmenten vergrössert. Die Segmentzahl ist z.T. an den Querfurchen der Rhachis bzw. an den Seitenfurchen erkennbar. Randstacheln können in der Verlängerung der Pleuren und damit entlang der ursprünglichen Segmentierung verlaufen, sie können aber auch sekundär und damit unabhängig von der Segmentation sein. Häufig ist am Pygidium ein hinterer Stachel oder ein unpaarer Fortsatz vorhanden. Bei Trilobiten, die sich gut einrollen konnten, hat das Pygidium häufig eine dem Kopfschild vergleichbare Grösse. Artikulationsmechanismen (Gruben und Höcker bzw. Furche und Kante) an Cephalon und Pygidium sorgten für eine entsprechend feste Fixierung bei der Einrollung. Der überwiegende Teil der Ventralseite sowie die Extremitäten sind weichkörperig und damit nur unter besonderen Umständen erhaltungsfähig. Zu beiden Seiten des Hypostoms inserieren die Antennen, gefolgt von undifferenzierten Spaltbeinen, von denen vier noch zur Kopfregion gehören. Die Gliedmassen dienen der Fortbewegung wie Laufen und Schwimmen, der Atmung und der Ernährung. Nachdem die ersten drei Beinpaare an Grösse zunehmen, verringert sie sich sukzessive nach hinten. Der Grundbauplan des Trilobitenbeins besteht aus einem Protopoditen, an dessen innerer Seite eine Gnathobase ansetzt. Vom Protopoditen spalten sich Exo- und Endopodit ab. Der Exopodit hat vermutlich respiratorische Funktion, während der Endopodit der Lokomotion dient.
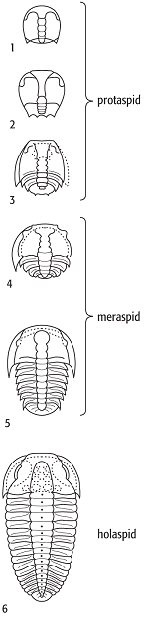
Die Ontogenie gliedert sich in drei Hauptphasen. Nach dem Schlüpfen befindet sich die Larve im sog. Protaspisstadium. Die Panzer sind ungeteilte ovale, mehr oder weniger stark gewölbte Schilde, die aber bereits die Duplikatur ausgebildet haben; die Ventralseite wird grösstenteils vom Hypostom bedeckt. Manchmal ist schon ein zentraler segmentierter Lobus als Andeutung der Glabella vorhanden. Im späteren Protaspis-Stadium entwickelt sich ein Protopygidium, das aber noch fest mit dem Kopfschild verwachsen ist. Im Meraspis-Stadium bildet sich ein Gelenk zwischen Kopf- und dem noch provisorischen Schwanzschild aus. Im weiteren Verlauf bilden sich die Thorakalsegmente. Jedes neue Segment entspricht meist einer Meraspis-Phase. Es kommt zu einer deutlichen Grössenzunahme, auch das Cephalon erhält jetzt die Adultmerkmale. Mit dem Holapsis-Stadium ist die endgültige Segmentzahl erreicht, die definitive Grösse wird durch weitere Häutungen erlangt (Abb. 3). Als Phaselus-Larven werden ovoide gewölbte Schalen interpretiert, die noch kleiner als das früheste Protaspisstadium sind.
Im Laufe ihrer ca. 250 Millionen Jahre währenden Existenz dokumentieren die Trilobiten eine grosse morphologische Diversität, ohne jedoch ihren Grundbauplan zu verändern. Obwohl meist adaptiver Natur, sind gewisse Merkmale Ausdruck evolutiver Trends. Man nutzt diese zur Rekonstruktion der Entwicklung und als Basis für eine Klassifikation. Zu diesen Merkmalen gehören u.a. die Gesichtsbzw. Häutungsnähte. Solche Häutungsnähte sind von keiner anderen Arthropodengruppe bekannt. Grundsätzlich unterscheidet man fünf verschiedene Typen: die propare, opisthopare, gonatopare, metapare und protopare oder hypopare Sutur. Ihr jeweiliger Verlauf ist aus Abb. 4 ersichtlich. Bei den Häutungsvorgängen platzt der Panzer entlang der oben genannten Suturen auf. Eine besondere Art der Häutung ist als Saltersche Einbettung bekannt und kommt bei den Phacopiden vor. Die Lebensweise der stenohalinen Trilobiten war in der Mehrzahl an den Boden gebunden, wo sie jedoch sehr unterschiedliche Nischen besiedelten. Harpiden und manche devonischen Scutelliden gelten als Riffbewohner (Riff). In Flachwasserkalken findet man im Ordovizium Illaeniden und Cheiruriden, im Silur z.B. Lichiden und Calymeniden, im Devon die Proetiden. Devonische Phacopiden dagegen kommen sowohl im kalkigen Flachwasserbereich als auch in anderen Fazies vor. In küstenfernen feinklastischen Sedimenten des Kambriums sind v.a. Oleniden vertreten. In der unterkarbonischen Kulmfazies (Kulm) kommen hauptsächlich kleinäugige bis blinde Formen vor, während in der Kohlenkalkfazies (Kohlenkalk) grossäugige Taxa dominieren. Die meisten Trilobiten waren an den Schelfbereich, d.h. bis ca. 200 m Wassertiefe, gebunden und ausserstande, den offenen Ozean zu überqueren. Daher lassen sie einen ausgeprägten Provinzialismus erkennen.
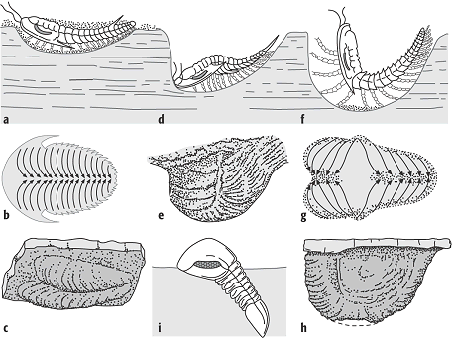
Die bei der Fortbewegung erzeugten Schreitfährten (Fährte) der Trilobiten geben z.B. Aufschluss über die Bewegungsrichtung, den Standort (auf oder im Sediment), die Art der Spur (z.B. Weide- oder Ruhespur) sowie über die Morphologie des Erzeugers. Ruhespuren (Rusophycus: Abb. 5) sind i.a. herzförmig, Kriechspuren (Cruziana) sind bandförmig. Je nach Verhalten der Tiere gibt es zwischen diesen beiden Spurentypen alle möglichen Übergänge.
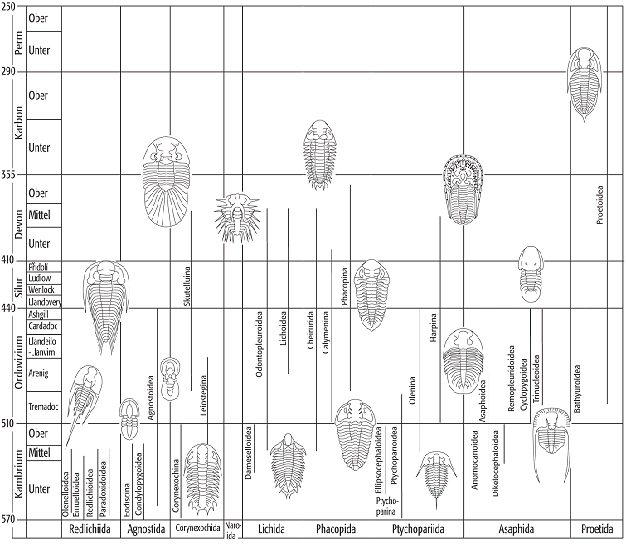
Systematisch unterscheidet man neun Trilobitenordnungen (Abb. 6): a) Redlichiida (Unter- bis Mittelkambrium): mit grossem, halbkreisförmigem Cephalon, zahlreichen, meist bestachelten Thorakalsegmenten und kleinem Pygidium. b) Agnostida (Unterkambrium bis Oberordovizium): mit fast gleichgrossem Kopf- und Schwanzschild und lediglich zwei Thorakalsegmenten, Formen meist blind, Lebensweise mit aufeinandergeklapptem Kopf- und Schwanzschild. c) Naraoiidae (Mittelkambrium, Burgess Shale): ohne Thorakalsegmente, Panzer unverkalkt. d) Corynexochida (Unterkambrium bis Mitteldevon): sehr heterogene Gruppe, Glabella parallel begrenzt oder nach vorn erweitert, opisthopare Sutur, 7-8 Thorkalsegmente, Hypostom z.T. mit Rostralplatte verschmolzen. e) Lichida (Mittelkambrium bis Mitteldevon): stark bestachelte Trilobiten und unter Umständen sehr grosse Formen mit z.T. tuberkuliertem Panzer, Glabella mit bis zu vier Glabellarfurchen, opisthopare Sutur, Grösse des Pygidiums je nach Untergruppe verschieden. f) Phacopida (Unterordovizium bis Unterdevon) mit vorherrschend proparen Suturen. Unterteilung in drei Unterordnungen: Cheirurina (mit nach vorn verbreiterter Glabella und vier Glabellarfurchen, 8-19 Thorakalsegmente, Pygidium bestachelt oder ausgefingertem Rand), Calymenina (mit nach vorn verschmälerter Glabella und gonatoparer Sutur, Thorax mit 11-13 Segmenten, Pygidium gerundet oder subdreieckig) und Phacopina (mit proparer Sutur, Glabella nach vorn verbreitert, ohne Rostralplatte, Thorax mit 11 Segmenten, Pygidium klein, gut entwickelter Einrollungsmechanismus). g) Ptychopariida (Unterkambrium bis Unterdevon): grosse paraphyletische Gruppe mit stark modifizierten Gruppen, typische Vertreter mit nach vorn verschmälerter Glabella und geraden Glabellarfurchen; grosser Thorax, aber kleines Pygidium. h) Asaphida (Unterkambrium bis Silur): Glabella undeutlich begrenzt, 8 Rumpfsegmente und grosses bis sehr grosses Pygidium, breiter Umschlag des Panzers auf Ventralseite. i) Proetida (Ordovizium bis Perm): grosse stark gewölbte Glabella, opisthopare Sutur, Cephalon mit Wangenstacheln, Rostralplatte nach hinten schmaler werdend, Thorax mit 8-10 Segmenten, Pygidium meist gefurcht und unbestachelt.
Über die Vorläufer der Trilobiten ist fast nichts bekannt, manche Autoren sehen Vertreter der Ediacara-Fauna oder Vendobionta als weichhäutige Vorfahren an. Die älteste Trilobitengruppe sind die Olenellina mit einem winzigen Pygidium. Gemeinsam mit den sekundär erblindeten Agnostida entwickeln sie eine gewisse Diversität im Kambrium. Hinzu kommen die Corynexochida und Ptychopariida, die aber auch nach dem Niedergang der Olenellina an der Wende Kambrium/Ordovizium weiterlebten. Die oberkambrische Fauna wird beherrscht von ptychopariiden Taxa. Mit Ausgang des Kambriums kam es zu einer grösseren Krise, in deren Verlauf die meisten der vergleichsweise unspezialisierten Formen ausstarben. Die genauen Gründe sind unbekannt, vermutlich spielten die oberkambrische marine Regression und das Auftreten räuberischer Cephalopoda auch eine Rolle. Nachdem im Tremadoc einige kurzlebige Formen auftauchten und wieder verschwanden, erschienen die ersten Vertreter der wichtigen und dominierenden ordovizischen Gruppen wie z.B. Phacopida, Asaphoida und Trinucleina. Diese ordovizischen Formen eroberten ganz neue Lebensbereiche, beispielsweise Riffe. Trilobitenfaunen des Silurs and Devons können leicht verwechselt werden, da sich viele Taxa überschneiden. Nur die Encrinuriden (Cheiruridae) reichen nicht sehr weit ins Devon hinein; sie sind ein typisches Element des Silurs. Die Mehrzahl der Trilobitengruppen stirbt im Mittel- bzw. Oberdevon aus. Nur die Proetiden überleben bis ins obere Perm. Die meisten von ihnen sind grossäugige Flachwasserbewohner, es gibt aber auch spezialisierte dünnschalige und blinde Formen des tieferen Wassers, die vom Oberdevon bis ins Kulm vertreten sind. Das Aussterben der letzten permischen Trilobiten hängt wahrscheinlich mit dem Absinken des Meeresspiegels zusammen, das auch viele andere Invertebratengruppen (Invertebraten) vorübergehend stark beeinflusst hat.
Grundsätzliche evolutionäre Errungenschaften beziehen sich auf das Entstehen neuer Augentypen, die Verbesserung der Einrollung sowie des Artikulationsmechanismus, der Wechsel von sehr kleinen zu grösseren Pygidien, die Ausbildung sehr starker Stacheln in einigen Gruppen sowie die Reduktion der Rostralplatte. Allerdings kam es seit Beginn des Ordoviziums zu keinen bedeutenden Änderungen mehr.
Der stratigraphische Nutzen der Trilobiten ist sehr gross; vor allem die Zonierung des Kambriums basiert fast ausschliesslich auf Trilobiten. Beschränkungen für ihre Anwendung liegen in ihrer fazieskontrollierten Verbreitung sowie in ihrem Provinzialismus. Im Ordovizium werden Ablagerungen des küstennahen Bereichs ebenfalls mit Hilfe von Trilobiten altersmässig eingestuft. Schwierigkeiten gibt es allerdings mit der zeitlichen Korrelation verschiedener Riffablagerungen und zeitgleicher gebankter Carbonate oder Kalkschlammfazies (Fazies). Für die stratigraphische Gliederung des Silurs und Devons haben sie lediglich eine gewisse lokale Bedeutung. Literatur: [1] Buchholz, A.(1997): Trilobiten mittelkambrischer Geschiebe aus Mecklenburg-Vorpommern (Norddeutschland). - Archiv für Geschiebekunde 4(2). [2] Fortey, R.A., Owens, R.M. (1999): The Trilobite Exoskeleton. In Savazzi, E. (ed.): Functional Morphology of the Invertebrate Skeleton. [3] Krueger, H.-H. (i. Dr.): Die Erratencrinurus Gruppe aus baltoskandischen Geschieben. - Palaeontographica A.
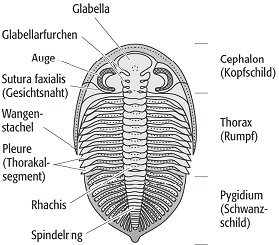 Trilobita 1: Dorsalansicht eines Trilobitenpanzers. Trilobita 1: Dorsalansicht eines Trilobitenpanzers.
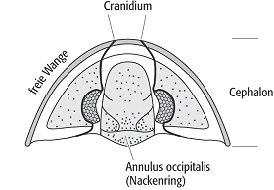 Trilobita 2: Merkmale eines Trilobitenkopfschildes. Trilobita 3: Ontogenese bei Sao hirsuta (Ptychopariida). Trilobita 2: Merkmale eines Trilobitenkopfschildes. Trilobita 3: Ontogenese bei Sao hirsuta (Ptychopariida).
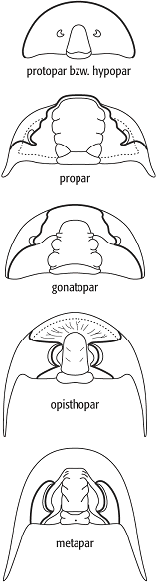 Trilobita 4: Gesichts- oder Häutungsnähte. Trilobita 5: Ruhespuren (Rusophycus) von Trilobiten: A-C) flaches Einscharren; F-H) Ausscharren einer Grube. Trilobita 4: Gesichts- oder Häutungsnähte. Trilobita 5: Ruhespuren (Rusophycus) von Trilobiten: A-C) flaches Einscharren; F-H) Ausscharren einer Grube.
 |
|
