| |
Spurenfossilien können als „versteinertes Verhalten” charakterisiert werden. Im Unterschied zu den Körperfossilien wird ein Organismus als Spurenfossil nicht mit seinen Hartteilen (dem Skelett), als kohliger Film oder als Abdruck überliefert, sondern über seine Aktivität. Es handelt sich um echte Fossilien, wenn man deren allgemeine Definition als physikalische Nachweise früherer Lebensformen zugrundelegt. Spurenfossilien stehen im Schnittfeld von Biologie und Sedimentologie und werden von der Ichnologie behandelt. Neben ihrer Fossilnatur sind sie gleichermassen auch Sedimentstrukturen, dürfen aber nicht mit den rein physikalischen Marken verwechselt werden: Selbst wenn ein Hartteil (z.B. Gehäuse, Zweig) eines toten Organismus diffuse Sedimentstrukturen wie Roll- oder Stossmarken verursacht, sind dies keine Zeugnisse seines Verhaltens zu Lebzeiten, also auch keine Spurenfossilien.
Spurenfossilien umfassen vor allem Belege der Fortbewegung und Ruhe von Tieren sowie Belege des Nahrungserwerbs und der Bodendurchwurzelung. Über die Abgrenzung zu Körperfossilien bzw. den Erzeugern der Spurenfossilien herrscht in Einzelfällen manchmal Unsicherheit. Keine Spurenfossilien, obwohl oft dafür gehalten, sind alle Strukturen, die von Organismen mittels Drüsen abgeschieden werden, also z.B. Eier, Perlen, Gallensteine oder die Schleimmatten der Blaubakterien, die als Stromatolithen fossil werden. Auch die teilweise zu den Aedificichnia (s.u.) gerechneten Spinnennetze gehören nicht hierher, sondern zu den Körperfossilien, ebensowenig wie zusammengeklebte Röhren von Würmern, Insekten oder Einzellern. Bei schlechter Erhaltung eines Spurenfossils kann seine Unterscheidung von Pseudofossilien schwierig sein; in vielen Fällen hilft jedoch die Kenntnis von Anatomie und Verhalten möglicher Erzeugergruppen ebenso wie das genaue Studium der Einbettungsumstände.
Von allen produzierten Spuren wird nur ein sehr geringer Prozentsatz im Gestein überliefert. Dafür muss die betreffende Spur die Zone aktiver Bioturbation verlassen. Dies kann z.B. geschehen, wenn plötzliche Schüttungen eines Sedimentes (oft anderer Korngrösse) eintreten, oder wenn ein Organismus so tief gräbt bzw. bohrt, dass seine Spur nicht mehr von nachfolgenden überprägt wird. Aus diesen Gründen sind zwei Überlieferungssituationen überproportional repräsentiert: Fressbaue und Kultivierungsspuren (s.u.) tiefgrabender Erzeuger wegen ihrer Position im Stockwerkbau und ihrer von der Gesteinsumgebung abweichenden Verfüllung sowie Weidespuren und Kriechspuren an der Basis von Trübeströmen (Turbidit). Für Fälle wie den letztgenannten, in denen eine Wechsellagerung von Gesteinen unterschiedlicher Korngrösse gegeben ist, wurden zwei nebeneinander gebräuchliche, lagebezogene Begriffssysteme entwickelt, die sich jeweils auf die grobkörnigere Bank beziehen (Abb. 1). Diese Klassifikation deckt aber nur einen Teil der Bioturbationsphänomene ab. Wichtiger als die beschreibende Bezeichnung ist zunächst, sich vor Augen zu führen, ob das jeweilige Fossil dem Aussehen des Baues zu Lebzeiten des Erzeugers entspricht; hier können bei kumulativen Spurenfossilien erhebliche Unterschiede bestehen (Abb. 2). Auch gilt es zu beachten, dass sich bei veränderter Stabilität des Sediments Spuren sehr unterschiedlicher Bildungsart und -zeit im gleichen Horizont befinden können. So sind z.B. Bohrspuren neben Grabgängen durch zwischenzeitliche Zementation des Substrates möglich, ihre Lage relativ zu ihrer Besiedlungsfläche weicht aber stark voneinander ab.
Das Nichterkennen des Erhaltungstyps von Spurenfossilien kann gravierende Folgen haben: Beispielsweise wurde auf der Basis von Unterfährten (s.u.) gefolgert, dass manche Dinosaurier treibend im Wasser lebten – tatsächlich waren nur die Trittsiegel der Hände bei einer Unterfährte nicht mehr erhalten. In einer ähnlichen Situation wurden unsinnig hohe Bewegungsgeschwindigkeiten von mehr als 50 km/h gefolgert; beides schwer auszurottende Irrtümer über die Biologie der Dinosaurier.
Eine allgemein übliche Einteilung in Grossgruppen existiert derzeit nicht, da einzelne Spuren verschiedenen Kategorien angehören können. Hier seien folgende vier Gruppen unterschieden: Spuren in Weichgrund (Bioturbation), Spuren in Hartsubstrat (Bioerosion) sowie die vom Substrat unabhängige Kotspuren (Koprolithen bzw. Faecichnia) und Wurzelspuren (Rhizolithen). Nur auf die erste Gruppe wird an dieser Stelle näher eingegangen (Abb. 3).
a) Aedificichnia, Bautenspuren (von lat. aedificium=Gebäude und griech. ichnos=Spur): Strukturen oberhalb des Substrates (also Bauten im Unterschied zu Bauen), die von den Erzeugern aus verkitteten Fremdpartikeln (Sediment, Holz) errichtet werden. Enganliegende Röhren um Einzeltiere gehören nicht hierher, sondern nur sehr viel grössere Gebilde wie Termitenhügel oder die Kokonhüllen mancher Erdwespen. Auch die manchmal als Calichnia (Brutspuren) bezeichneten Strukturen für die Larvenaufzucht (z.B. Bienenwaben) sind wegen grosser Schwierigkeiten der Abgrenzung hierher zu rechnen, zumal sie extrem selten überliefert sind.
b) Agrichnia, Kultivierungsspuren, „Graphoglyptiden” (von lat. ager=Acker und griech. ichnos=Spur): Grabgänge von Tieren mit einer Mischernährung, die in ihren dauerhaft offenen Bauen einerseits Pilze oder Mikroorganismen züchten, um diese regelmässig abzuweiden, andererseits die Baue als Fallen für kleinere grabende Organismen benutzen; die Erzeuger waren also gleichzeitig Farmer und Trapper. Agrichnia zeigen immer einen geometrischen Aufbau in Form von Netzen, Spiralen, Mäandern etc. (Beispiele: Paleodictyon, Belorhaphe, Cosmorhaphe und die meisten Arten von Chondrites).
c) Cubichnia, Ruhespuren (von lat. cubare=liegen und griech. ichnos=Spur): Spurenfossilien, die durch vagile Tiere während ihrer Ruhephasen verursacht wurden. Sie sind in die Substratoberfläche eingemuldet und geben den Körperumriss des Erzeugers etwas verwaschen wieder (Beispiele: Asteriacites, Lockeia, Rusophycus).
d) Domichnia, Wohnbaue (von lat. domus=Haus und griech. ichnos=Spur): Spurenfossilien, die hemisessilen, filtrierenden oder angelnden Tieren als Dauerwohnung gedient haben. Es handelt sich meist um senkrechte Röhren (gestreckt oder U-förmig), die in gut durchlüftetem Milieu angelegt wurden (Beispiele: Diplocraterion, Ophiomorpha, Skolithos); auch fast alle Bohrgänge gehören hierher.
e) Equilibrichnia,Ausgleichsspuren(von lat. equilibrium=Gleichgewicht und griech. ichnos=Spur):Spurenfossilien von grabenden Tieren in enger Verwandtschaft zu den Fugichnia, doch durch regelmässige Anpassung an höhere und niedrigere Substratoberflächen entstanden. Im Querschnitt der vertikalen Struktur ist die Sedimentschichtung deutlich erkennbar; Ichnotaxa sind dafür bisher nicht vergeben worden.
f) Fodinichnia, Fressbaue (von lat. fodina=Bergwerk und griech. ichnos=Spur): Spurenfossilien, die von tiefgrabenden, sedimentfressenden Tieren verursacht wurden. Für die beste Nutzung des organischen Materiales aus dem sauerstoffarmen Sediment sind die Baue geometrisch planvoll (radial, U-förmig etc.) angelegt (Beispiele: Rhizocorallium, Thalassinoides und viele Arten von Zoophycos). g) Fugichnia, Fluchtspuren (von lat. fugare=fliehen und griech. ichnos=Spur): Spurenfossilien, die von grabenden Tieren nach plötzlicher Verschüttung verursacht wurden. Vor allem Muscheln, Seesterne und diverse „Würmer” befreien sich durch schnelle Verlagerung ihrer Position nach oben, was im Sediment ein vertikales Band schlieriger Struktur hinterlässt. Übergänge zu Equilibrichnia, Cubichnia und Domichnia sind häufig; Vertreter dieser Spurengruppe wurden bisher nicht benannt. h) Pascichnia, Weidespuren (von lat. pascere=weiden und griech. ichnos=Spur): Spurenfossilien, die an der Oberfläche durch pflügende, sedimentfressende oder algenabweidende Tiere verursacht wurden. Bei nur mässiger Sauerstoffversorgung wurde sich bodennah ansammelnder Detritus nicht sofort zersetzt, so dass die Erzeuger der Pascichnia darauf ihre spezialisierte Ernährung aufbauen konnten. Es handelt sich um unverzweigte Bandstrukturen, die sich oftmals mäandrierend zu engstehenden, ornamentalen Mustern zusammenschliessen (Beispiele: Helminthopsis, Nereites, Phycosiphon).
i) Praedichnia, Raubspuren, Frassspuren (von lat. praedare=rauben und griech. ichnos=Spur): Spurenfossilien, die durch Räuber an ihrer Beute verursacht wurden. Hierzu zählen die typischen Bohrlöcher (Bioerosion) von zwei Schneckenfamilien und von Kraken, die Bissspuren von Raubtieren an Knochen und die von Krabben zerknackten Weichtiergehäuse. Der ältere Name Mordichnia hat sich nicht durchsetzen können. Auch die Spuren des Insektenfrasses an Pflanzen gehören hierher: Schmetterlinge, Käfer, Heuschrecken u.a. hinterlassen an Blättern gezackte Ränder, Löcher und die sog. Minen (Beispiele: Phagophytichnus, Oichnus).
j) Repichnia, Kriechspuren bzw. Fährten (von lat. repere=kriechen und griech. ichnos=Spur): Spurenfossilien, die durch die Bewegung auf dem Substrat verursacht wurden. Anzeichen von Fressakten sind nicht erkennbar. In aller Regel handelt es sich um (doppel)bandförmige, lineare oder weit geschwungene Strukturen; auch die von schwimmenden Tieren bei Bodenberührung hinterlassenen, geschlängelten Spuren (Natichnia) werden hierzu gezählt (Beispiele: Cruziana, Diplichnites, Undichnus). Eine Abtrennung der reinen Trittsiegel-Fährten als Cursichnia hat sich nicht durchsetzen können.
Wichtig bei der Benennung (Ichnotaxonomie) ist: Spurenfossilien stehen in keiner eindeutigen Beziehung zu ihren Erzeugern: Unterschiedliche Organismen können unter gleichen Bedingungen die gleiche Spur erzeugen und mehrere Erzeuger können bei der Bildung einer gemeinsamen Spur zusammenwirken. Auch kann ein und derselbe Organismus verschiedene Spuren verursachen, wenn sich sein Verhalten aufgrund der Art des Substrates ändert. Dadurch kann die gleiche Spur in unterschiedlichen Substraten anders aussehen, ohne dass hierfür ein spezieller Name erforderlich wäre. Auch bei gleichbleibenden Substrateigenschaften kann sich das Verhalten eines Erzeugers ändern, was zu ineinander übergehenden Spuren führt, die dann getrennt benannt werden müssen. Ein spezielles Problem stellen die Unterfährten von Wirbeltieren dar: Häufig drückt sich eine Extremität soweit in den Untergrund ein, dass ihr Umriss auf mehreren Schichtflächen erkennbar bleibt, allerdings um so undeutlicher und detailärmer, je weiter die Fläche von der tatsächlich betretenen Oberfläche entfernt ist (Abb. 4). Diese Unterfährten dürfen nicht benannt werden, da sie eine andere Fährtenform nur vortäuschen. Aus den obigen Gründen muss man vermeiden, eine Spur mit ihrem Erzeuger gleichzusetzen, selbst wenn dieser genau bekannt sein sollte. Spurenfossilien tragen vielmehr eigene Namen, die dem Internationalen Codex für Zoologische Nomenklatur (ICZN) unterliegen, also wie wissenschaftliche Namen von Tieren behandelt werden. Die Namen von Spurenfossilien, die Ichnotaxa, sind daher zweiteilig wie überall in der Biologie, d.h. sie bestehen aus grossgeschriebenen Gattungsnamen (Ichnogenus) und kleingeschriebenen Artnamen (Ichnospecies). Die Benennung eines konkreten Spurenfossils erfolgt im Rahmen der Ichnotaxonomie, derjenigen Teildisziplin der Ichnologie, die die Bestimmungskriterien festlegt. Als Kriterien, die sog. Ichnotaxobasen, können unterschiedliche Merkmale fungieren, die je nach Typ des Spurenfossils eingesetzt werden. Dazu zählen die generelle Form, die interne Struktur, ggf. der Bau der Wandung, die Skulpturierung der Aussenseite, die Orientierung zum Substrat, der Typ des Substrates und in seltenen Fällen die Grösse. Für Fährten sind wichtig die Symmetrie, der Abstand und das Wiederholungsmuster der einzelnen Trittsiegel sowie Breite und Durchgängigkeit der Fährte. Biologie der Erzeuger, Einbettungseffekte, Unterschiede in der räumlichen oder zeitlichen Verbreitung u.a. dürfen in keinem Fall verwendet werden.
Im Bereich der Meeressedimente erkennt man, dass mit dem Aufkommen der Trilobiten im unteren Kambrium die Bioturbationsintensität ebenso sprunghaft ansteigt wie die maximale Tiefe von Grabgängen: Die sich von schwebenden Kleintieren ernährenden Suspensionsfresser gruben sich zu dieser Zeit in 0-6 cm Tiefe, im darauffolgenden Ordovizium aber zwischen 6 und 12 cm. Das Einsetzen der typisch paläozoischen Fauna im mittleren Ordovizium hatte eine nochmals verstärkte Durchwühlung der Sedimente zur Folge, und bis zum Karbon waren die grabenden Suspensionsfresser schon in Tiefen von 12-100 cm zu finden. Hier drücken sich biologische Wechselwirkungen wie Konkurrenzvermeidung und Räuberdruck aus. Auch lässt sich der von diversen Gruppen der Körperfossilien bekannte Trend nachvollziehen, dass neue Baupläne in Küstennähe entstehen und sich nach einiger Zeit ins tiefere Wasser ausbreiten oder verlagern (bzw. abgedrängt werden). Keinen Niederschlag in der Überlieferung der Spurenfossilien finden dagegen die grossen Krisen der Geschichte des Lebens im Devon und am Ende von Perm und Kreide. Ab der mittleren Kreide ist vielmehr eine stark zunehmende Häufigkeit und Vielfalt der Tiefsee-Weidespuren festzustellen, die offenbar auf das Aufkommen der Bedecktsamer zurückgeht. Deren enormer Umsatz an Biomasse (Laubfall!) hatte Auswirkungen bis ins Meer, wo Detritusfresser wesentlich bessere Nahrungsgrundlagen fanden als zuvor. Literatur: [1] BOTTJER, D.J. & DROSER, M.L. (1992): Paleoenvironmental patterns of biogenic sedimentary structures; in: MAPLES, C.G. & WEST, R.R. (Hrsg.): Trace fossils. - Paleontol. Soc. Short Courses in Paleontol. 5: 130-144. Knoxville. [2] BROMLEY, R.G. (1999): Spurenfossilien. – Berlin/Heidelberg. [3] EKDALE, A.A., BROMLEY, R.G. & PEMBER-TON, S.G. (1984): Ichnology - the use of trace fossils in sedimentology and stratigraphy. – Soc. Econ. Paleontol. Mineral. Short Course 15. Tulsa. [4] SIMPSON, S. (1975): Classification of trace fossils; in FREY, R.W. (Hrsg.): The study of trace fossils: 39-54. Berlin/Heidelberg/New York.
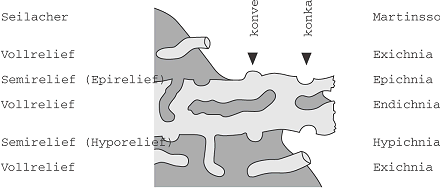 Spurenfossilien 1: Vergleich der lagebezogenen Einteilung von Spurenfossilien in einer Wechsellagerung. Spurenfossilien 1: Vergleich der lagebezogenen Einteilung von Spurenfossilien in einer Wechsellagerung.
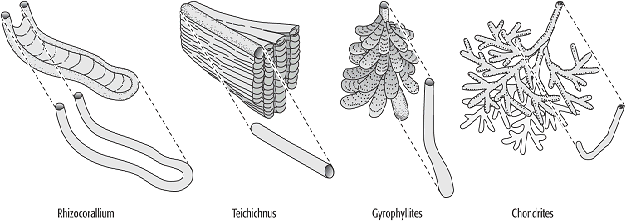 Spurenfossilien 2: Unterschiede zwischen den Bauen zu Lebzeiten der Erzeuger und den im Gestein erkennbaren Spurenfossilien. Spurenfossilien 2: Unterschiede zwischen den Bauen zu Lebzeiten der Erzeuger und den im Gestein erkennbaren Spurenfossilien.
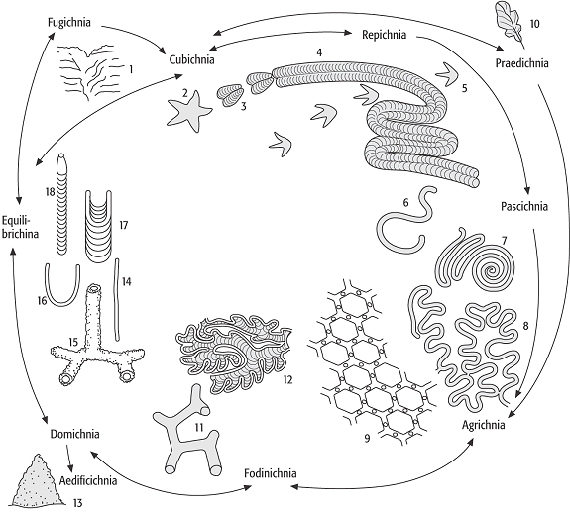 Spurenfossilien 3: Einteilung der Spurenfossilien nach dem Verhalten ihrer Erzeuger: 1) Fluchtspur einer Muschel, 2) Ruhespur (Asteriacites) eines Seesterns, 3) Ruhespur (Rusophycus) eines Gliedertiers, 4) Kriechspur (Cruziana) eines Gliedertiers, 5) Fährte eines zweibeinigen Wirbeltiers, 6) Weidespur Planolites, 7) Weidespur Helminthoida, 8) Kultivierungsspur Cosmorhaphe, 9) Kultivierungsspur Paleodictyon, 10) Frassspur (Phagophytichnus) eines Insektes, 11) Wohn/Fressbau (Thalassinoides) eines Krebses, 12) Fressbau Phycosiphon, 13) Termitenhügel als Bautenspur, 14) Wohnröhre (Skolithos) eines Wurmes, 15) Wohnbau (Ophiomorpha) eines Krebses, 16) Wohnröhre (Arenicolites) eines Wurmes, 17) Ausgleichsspur (Diplocraterion) eines Wurmes, 18) Ausgleichsspur einer Muschel. Spurenfossilien 3: Einteilung der Spurenfossilien nach dem Verhalten ihrer Erzeuger: 1) Fluchtspur einer Muschel, 2) Ruhespur (Asteriacites) eines Seesterns, 3) Ruhespur (Rusophycus) eines Gliedertiers, 4) Kriechspur (Cruziana) eines Gliedertiers, 5) Fährte eines zweibeinigen Wirbeltiers, 6) Weidespur Planolites, 7) Weidespur Helminthoida, 8) Kultivierungsspur Cosmorhaphe, 9) Kultivierungsspur Paleodictyon, 10) Frassspur (Phagophytichnus) eines Insektes, 11) Wohn/Fressbau (Thalassinoides) eines Krebses, 12) Fressbau Phycosiphon, 13) Termitenhügel als Bautenspur, 14) Wohnröhre (Skolithos) eines Wurmes, 15) Wohnbau (Ophiomorpha) eines Krebses, 16) Wohnröhre (Arenicolites) eines Wurmes, 17) Ausgleichsspur (Diplocraterion) eines Wurmes, 18) Ausgleichsspur einer Muschel.
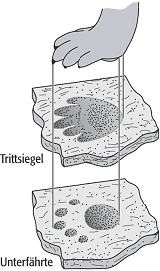 Spurenfossilien 4: Unterfährten und ihre Beziehung zu Fährten. Spurenfossilien 4: Unterfährten und ihre Beziehung zu Fährten. |
|
