| |
Porifera, aktiv filtrierende, vielzellige Tiere. Sie zeigen verwandtschaftliche Beziehungen zu kolonial organisierten, einzelligen Eukaryota und den Eumetazoa. Die rezenten Schwämme sind weitestgehend marin, es sind aber auch Vertreter aus dem Süsswasser bekannt. Wenige rezente Schwämme werden kommerziell genutzt (Schwammfischerei) und einige sind potentielle Lieferanten pharmazeutisch verwertbarer Naturstoffe. Physiologisch entsprechen die Schwämme einer natürlichen Kläranlage, indem sie gelöste und partikuläre organische Substanz über einen aktiven Filterapparat verwerten. Sie sind deshalb wichtig zur Rekonstruktion fossiler Nahrungsnetze und der Abschätzung ozeanographischer Rahmenbedingungen (z.B. Gehalte suspendierter Nahrung, Strömungsmuster). Fossile Schwämme sind häufig in Biostromen, riffähnlichen Biohermen und in regelrechten Schwammriffen zu finden. Als Riffbewohner sind die Schwämme in fossilen wie rezenten Korallenriffen stets vertreten. Die Gesamtgruppe weist ein stark unterschiedliches Mass der Skelettisierung auf, entsprechend unterschiedlich und selektiv verläuft die Fossilisation im geologischen Bericht. Der Grundbauplan und die allgemeine Physiologie eines Schwammes ist in Abbildung 1 dargestellt. Durch koordinierten Geisselschlag der in Kammern organisierten Kragengeisselzellen (Choanocyten, Choanocytenkammern, Choanoderm) wird eine Pumpwirkung erzielt. Dabei wird Wasser aus der Umgebung durch Einlassporen (Ostien) in den Schwammkörper geleitet, filtriert und durch kaminartige zentrale Öffnungen (Oscula) wieder ausgetrieben. Bei den meisten Schwämmen ist dieses Kanalsystem durch Plattenzellen (Pinacocyten) begrenzt, die zu einer Deckzellschicht organisiert sind (Pinacoderm). Porentragende Zellen (Porocyten) können im Pinacoderm eingelassen sein. Das Pinacoderm umschliesst das innere Schwammgewebe (Mesohyl), in dem sich eine Vielzahl von Zelltypen befinden. Neben den Stammzellen (Archaeocyten) kommen u.a. verschiedene skelettsubstanzbildende Zellen (Lophocyten, Scleroblasten, Spongioblasten), kontraktile Zellen (Collencyten), Speicherzellen (Trophocyten) und Keimzellen (Oogonien, Spermatogonien) vor. Aus den Archaeocyten können alle anderen Zelltypen eines Schwammes hervorgehen (Omnipotenz). Viele Schwämme führen in ihrem Mesohyl eine Vielzahl verschiedenster Bakterien, die zumindest teilweise zum Schwamm in symbiotischer Beziehung stehen. Die Reproduktion bei Schwämmen erfolgt sexuell (häufig Zwitter) und asexuell durch Knospung. Dauerstadien (Gemmulae) und Regenerationsfähigkeit aus Fragmenten sind nachgewiesen (Aktivierung der Archaeocyten). Die sexuelle Reproduktion führt über ein planktisches Larvalstadium und der Larvenansiedelung zum sessilen Jungtier.
Die typischen Merkmale der Eumetazoen (Vielzelligkeit mit Stammzellen, sexuelle Reproduktion mit planktischen Larvalstadien, Kollagensynthese und Zellgewebe) sind zwar vorhanden, doch fehlen die für alle anderen Metazoen kennzeichnenden Merkmale eines echten Epithelgewebes. Deshalb werden die Schwämme häufig als Vorläufer der Eumetazoen interpretiert und als sogenannte Parazoa bezeichnet. Alternativ dazu wird aufgrund der spezifischen Merkmale eines Schwammes (choanodermal organisierter, aktiv filtrierender Vielzeller) eine mehrfache und unabhängige Evolution hin zur Vielzelligkeit angenommen (z.B. über kolonial organisierte Choanoflagellaten). Folgt man dieser Vorstellung, stehen die Schwämme auf der gleichen phylogenetischen Rangstufe wie alle übrigen Metazoen zusammen.
Die Schwämme weisen zwei grundsätzlich verschiedene Typen der Skelettbildung auf. Zum Aufspannen des Schwammkörpers dient ein Stützskelett. Einige Schwämme bilden zusätzlich einen Kalksockel zwischen Substrat und Weichkörper aus (sekundäres Basalskelett). Das Stützskelett besteht aus organischer Substanz (Kollagen-, Sponginfasern) und ist häufig mit Schwammnadeln aus einer mineralischen Komponente (Skelettopal, SiO2, oder Mg-Calcit, CaCO3 mit ca. 10 Mol-% MgCO3) kombiniert. Neben der unterschiedlichen Mineralogie weisen diese Schwammnadeln (Skleren oder Spiculae) unterschiedliche Geometrien, eine hohe Variation der räumlichen Anordnung (Sklettarchitektur) und verschiedene Vernetzungsgrade auf. Zudem sind bei vielen Schwämmen unterschiedliche Sklerentypen kombiniert, und es können zwei diskrete Grössenordnungen auftreten (Mega- bzw. Mikroskleren jeweils unterschiedlicher Entstehung). All diese, grösstenteils auch für die Paläontologie verwertbaren Merkmale dienen zusammen mit histologischen und ontogenetischen Befunden der weiteren taxonomischen Untergliederung der Schwämme. Gut begründete, geschlossene Abstammungsgemeinschaften (monophyletische Gruppen) sind die Hexactinellida, die Calcarea und die Demospongea. Die phylogenetischen Zusammenhänge zwischen diesen Gruppen sind aber weiterhin in Diskussion. Die Hexactinellida (Glasschwämme) weisen neben einer besonderen Weichkörperorganisation (Choanosyncytium) auch einen spezifischen Sklerentypus auf. Die Grundform entspricht einem regelmässigen Sechsstrahler (Hexactin) aus SiO2 (Abb. 2). Diese Kieselskleren werden intrazellulär gebildet. Paläontologisch bedeutsam sind die unterschiedlichen Vernetzungsgrade des Sklerenskelettes. Man unterscheidet zwischen a) locker angeordneten Skleren (lyssakines Skelett, Lyssakinosa), b) durch eine weitere SiO2-Hülle miteinander verkittete Skleren (dyktionales Skelett, Hexactinosa) und c) durch zentrale Querstreben stabilisierte Skleren (lychniskoses Skelett,
Lychniscosa). Die ältesten Nachweise der Hexactinellida sind Lyssakinosa aus dem Neoproterozoikum (Vendium, ca. 545 Mio. Jahre). Die Hexactinosa erscheinen im Devon (Oberdevon, Frasne), die Lychniscosa im mittleren Jura (Dogger). Die heutigen Hexactinellida leben ausschliesslich in marinem Milieu, vorzugsweise in der Tiefsee, und sind meist durch spezielle Skleren mit Weichböden verwurzelt. Die Häufigkeit und Verbreitung der fossilen Hexactinellida ist starken Schwankungen unterworfen. Neben den mit den heutigen Verhältnissen vergleichbaren Vorkommen sind regelrechte Hexactinelliden-Rasen und durch Hexactinelliden geprägte mud mounds bekannt. Ausserordentliche Vorkommen befinden sich im Burgess Shale (Kambrium), im Jura (Malm) z.B. der schwäbischen und fränkischen Alb und in der Kreide Niedersachsens und Westfalens (z.B. Coeloptychium-Schichten, Campan). In diesen Fällen sind grossräumige Schelfareale besiedelt worden. Unter Umständen handelt es sich deshalb bei den heutigen Vorkommen um in tiefere Gewässer verdrängte Gemeinschaften, eventuell auch um ein Relikt der quartären Vereisung.
Die Calcarea (Kalkschwämme) bilden extrazellulär unter Beteiligung mehrerer Skleroblasten Mg-calcitische Skleren (Abb. 3). Die Grundformen der Skleren entsprechen dem Triactin (regelmässiger Dreistrahler) und dem Octactin (regelmässiger Achtstrahler). Octactine sind nur von den auf das Paläozoikum beschränkten Heteractinida bekannt. Vom Triactin abgeleitet kommen sogenannte Stimmgabel-Triänen vor, die für das Taxon der Minchinellidae kennzeichnend sind. Einstrahlige Skleren (Monactine) können ebenfalls vorhanden sein. Häufig bestehen dicht verfilzte Sklerenzüge, die von einem kalkigen, sekundären Skelett umgeben sind. Dazu zählen die Pharetroniden und ein Vertreter der rhythmisch gekammerten Kalkskelette der Sphinctozoen. Die heutigen Kalkschwämme leben bevorzugt in tropischen Flachwassergebieten, kommen als Einzelfunde aber auch in der Tiefsee vor. Dies stimmt gut mit dem geologischen Bericht überein. Die Kalkschwämme sind ab dem Kambrium nachgewiesen. Die Pharetroniden sind v.a. in tropischen Ablagerungen des Mesozoikums (Jura und Kreide) zu finden, bilden aber nur vereinzelt eine hohe Besiedlungsdichte aus.
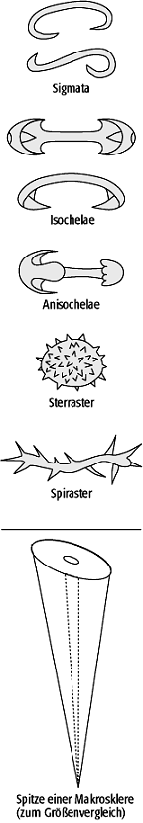
Die Demospongea nehmen den weitaus grössten Teil der heutigen und fossilen Schwammtaxa ein. Neben den marinen Formen, inklusive der Bohrschwämme (z.B. Cliona), gehören hierzu auch die Süsswasserschwämme (z.B. Ephydatia). Kennzeichnendes Merkmal der Demospongea ist die intrazelluläre Bildung von Kieselskleren mit der Grundform eines regelmässigen Vierstrahlers (Tetractin, Calthrops). Diese Grundform wird z.T. erheblich variiert, so dass insgesamt eine grosse Formenvielfalt der Skleren vorliegt (Abb. 4). Weitere Merkmale beziehen sich auf die Sklerenarchitektur sowie das Vorkommen und den Typus von Mikroskleren. Die Sklerenbildung kann auch vollständig reduziert sein, so dass ausschliesslich organische Stützskelette vorliegen (z.B. bei dem Badeschwamm Spongia officinalis). Diese Schwämme können jedoch kaum fossilisiert werden. Innerhalb der Demospongea sind die Taxa Homoscleromorpha, Ceractinomorpha und Tetractinellida gut als Monophylum begründet. Bei den Homoscleromorpha existiert keine Differenzierung zwischen Mega- und Mikroskleren. Die Ceractinomorpha besitzen einstrahlige Megaskleren und Mikroskleren vom Sigma- oder Chela-Typus, die Tetractinellida haben tetractine Megaskleren und Mikroskleren vom Aster-Typus.
Der Grad der Sklerenvernetzung und die Beständigkeit bzw. Festigkeit des Sklerenskelettes bestimmt weitgehend das Fossilisationspotential und die Qualität der Erhaltung. Nichtrigide Demospongea sind nur in Ausnahmefällen im quasi-primären Zusammenhang erhalten. Diese Ausnahmefälle beziehen sich auf verborgene und geschützte Kleinhabitate, z.B. in den tieferen Zonen von Riffen oder in Bohrlöchern und absoluten Stillwasserzonen. Ein besonderes Beispiel hierfür sind Massenvorkommen von Skleren (Spikulite), die die fossilen Reste ehemaliger Demospongea-Matten repräsentieren. Die Regel allerdings ist, dass die Skleren nichtrigider Demospongea (analog zu den Lyssakinosa) nicht im primären Verband, sondern als isolierte und meist transportierte Skleren in Sedimentgesteinen vorliegen. Im Gegensatz zu den nichtrigiden Demospongea mit niedrigem Erhaltungspotential weisen die rigiden Demospongea ein hohes Erhaltungspotential auf und sind damit im geologischen Bericht deutlich überbewertet. Für diesen Typus stehen die Lithistida (Steinschwämme), eine polyphyletische Gruppe desmentragender Demospongea. Desmen sind unregelmässig bedornte und zerlappte Megaskleren, die sich gegenseitig über bedornte Endungen (Zygome) zu einem rigiden Sklerengerüst verbinden können. Dadurch sind die Lithistida häufig vollkörperlich erhalten und können sogar kurzzeitigen Transport überstehen.
Die vermutlich ältesten Demospongea stammen aus dem Neoproterozoikum (frühes Vendium, ca. 580 Mio. Jahre). Spikulite treten mehrfach auf und sind häufig an Zonen kühler und an Nährstoffen reicher Wässer gebunden (z.B. glaziale Episode im Grenzintervall Ordovizium zu Silur; z.T. auch im Flammenmergel der Unterkreide Mitteldeutschlands). In Biostromen und Biohermen des Phanerozoikums können die Demospongea (v.a. Lithistida) massgeblich beteiligt sein. Wichtige Vorkommen stammen aus dem Burgess Shale (Kambrium), dem Ordovizium und Silur (z.B. Ostkanada), dem Jura (Spanien, Portugal, Schwäbische und Fränkische Alb) und der Kreide (Spanien, Mitteldeutschland). Zusammen mit den Hexactinelliden bilden die Lithistida das prägende Element z.B. der jurassischen Schwamm-Bioherme (mud mounds). Im Vergleich zu den Hexactinelliden reichen die Demospongea fossil wie rezent bis in flachere Schelfgebiete und sind wesentlich in und um moderne Korallenriffe vertreten. Über das Verhältnis der Hexactinellida zu den Lithistida kann auch im geologischen Bericht eine grobe Abschätzung der relativen Tiefenlage der Ablagerungsräume erfolgen. In feinkörnigen Carbonatsedimenten werden die Demospongea (z.T. auch die Hexactinellida) diagenetisch stark überprägt. Zu dieser Fossildiagenese kann eine umfassende Calcifizierung, die Auflösung des Skelettopals, Calcitzementation sowie eine lokale Pyritisierung und sekundäre Verkieselung gehören.
Vor allem innerhalb der verschiedenen Taxa der Demospongea (vereinzelt auch bei den Calcarea) gibt es Schwämme, die zusätzlich zu ihrem primären Stützskelett ein sekundäres, kalkiges Basalskelett ausbilden (aus Calcit oder Aragonit). In früheren Jahren als monophyletische Gruppe interpretiert (Klasse Sclerospongia), ist mittlerweile die Polyphylie dieser Gruppe mehrfach unabhängig voneinander belegt worden. Die Vertreter dieser besonderen Skelettisierung werden heute als coralline Schwämme zusammengefasst. Das Fossilisationspotential des kalkigen Basalskelettes ist besonders hoch.
Morphologisch können im wesentlichen fünf Basalskelett-Typen unterschieden werden: a) Der Krusten-Typus entspricht einem einfachen Kalksockel ohne vertikale Differenzierung. b) Der chaetetide Typus (Abb. 5) weist ein durch Zwischenböden (Tabulae) untergliedertes Röhrensystem (Tuben) auf (Chaetetida). c) Stromatoporoide Basalskelette (Stromatoporen; Abb. 6) entsprechen offenen Netzwerken mit horizontal (laminar) und vertikal ausgerichteten Elementen (Pfeiler). Häufig sind blasige Strukturen ausgebildet (Vesikulae) und es bestehen durchhaltende Röhren eines Exkretionsystems, das an der Oberfläche auf Erhebungen ein sternförmiges Muster ausbildet (Astrorhizen). d) Sphinctozoide bzw. thalamide Typen sind rhythmisch gekammerte bzw. segmentierte Basalskelette meist zylindrischer Wuchsform. e) Der ausgestorbene Archaeocyathen-Typus (Unter- bis Mittelkambrium) entspricht im wesentlichen einem doppelwandigen, mit Poren durchsetztem Kegel (Archaeocyathida).
Die heutigen corallinen Schwämme sind vor allem aus submarinen Höhlen, verborgenen (kryptischen) Riffhabitaten und tieferen Vorriffzonen bekannt (Mittelmeer, Karibik, Indopazifik). Die archaeocyathiden Schwämme waren am Bau der durch verkalkende Mikroorganismen geprägten Riffe des Kambriums beteiligt, meist jedoch auf kryptische Habitate limitiert (sogenannte Archaeocyathen-Riffe, z.B. Sierra Morena, Spanien). Stromatoporoide Schwämme waren kennzeichnende Organismen v.a. devonischer Riffe und chaetetide Schwämme waren im Karbon wesentlich an der Bildung von Biostromen und Biohermen beteiligt. In grossen Teilen von Perm und Trias gab es von sphinctozoiden Demospongea dominierte Riffstrukturen (z.B. Djebel Tebaga in Tunesien, Kalkalpen). Ab der Unterkreide bestanden mit den heutigen Vorkommen coralliner Schwämme gut vergleichbare Verhältnisse (weitgehend identische Gattungen, analoge Habitate). Literatur: [1] Ax, P. (1995): Das System der Metazoa I. - Stuttgart. [2] Boardman, R.S., Cheetham,
A.H. & Rowell, A.J. (Hrsg.) (1987): Fossil Invertebrates. - Oxford. [3] Reitner, J. & Keupp (Hrsg.) (1991): Fossil and recent sponges. - Berlin. [4] van Soest, R.W.M., Kempen, T.M.G. & Braekman,
J.C. (Hrsg.) (1994): Sponges in time and space. - Rotterdam. [5] Weissenfels, N. (1989): Biologie und mikroskopische Anatomie der Süsswasserschwämme (Spongillidae). - Stuttgart. [6] Westheide, W. & Rieger, R. (Hrsg.)(1996): Spezielle Zoologie, Teil 1: Einzeller und Wirbellose Tiere. - Stuttgart.
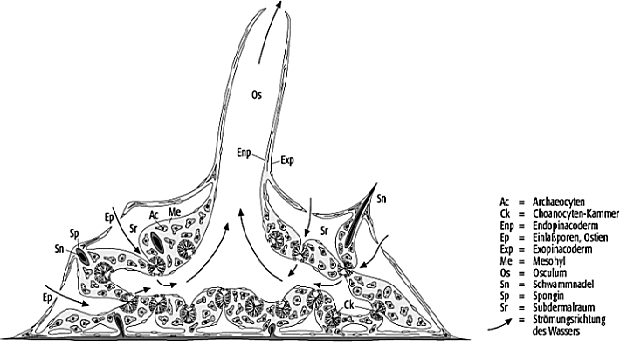 Schwämme 1: Grundbauplan eines Schwammes am Beispiel eines Süsswasserschwammes (Spongillidae, Demospongea). Schwämme 1: Grundbauplan eines Schwammes am Beispiel eines Süsswasserschwammes (Spongillidae, Demospongea).
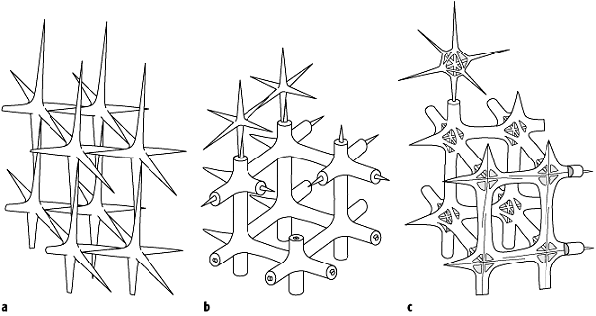 Schwämme 2: Regelmässige Sechsstrahler (Hexactine) bilden die Grundform der Megaskleren bei den Hexactinellida. Die Art der Sklerenvernetzung führt zur Unterscheidung von a) Lyssakinosa, b) Hexactinosa und c) Lychniscosa. Schwämme 2: Regelmässige Sechsstrahler (Hexactine) bilden die Grundform der Megaskleren bei den Hexactinellida. Die Art der Sklerenvernetzung führt zur Unterscheidung von a) Lyssakinosa, b) Hexactinosa und c) Lychniscosa.
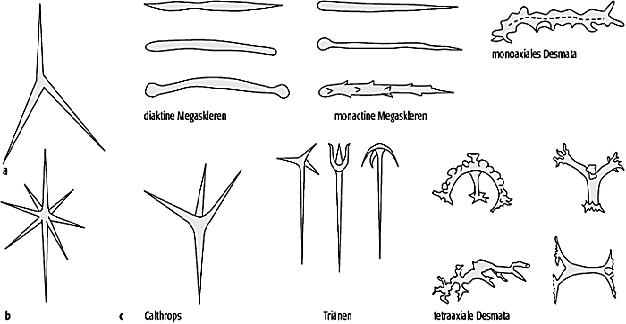 Schwämme 3: Megaskleren der Calcarea (a=triactine Kalksklere, b=octactine Kalksklere) und der Demospongea (c). Schwämme 4: Mikroskleren der Demospongea. Schwämme 3: Megaskleren der Calcarea (a=triactine Kalksklere, b=octactine Kalksklere) und der Demospongea (c). Schwämme 4: Mikroskleren der Demospongea.
 Schwämme 5: coralliner Schwamm mit chaetetidem Basalskelett: Acanthochaetetes aus der Unterkreide Nordspaniens (Gesteinsdünnschliff). Schwämme 5: coralliner Schwamm mit chaetetidem Basalskelett: Acanthochaetetes aus der Unterkreide Nordspaniens (Gesteinsdünnschliff).
 Schwämme 6: coralliner Schwamm mit stromatoporoidem Basalskelett: Actinostromaria aus der Unterkreide Nordspaniens (Gesteinsdünnschliff). Deutlich zu sehen ist das nach oben sternförmig zulaufende Exkretionsystem (Astrorhizenkanäle). Schwämme 6: coralliner Schwamm mit stromatoporoidem Basalskelett: Actinostromaria aus der Unterkreide Nordspaniens (Gesteinsdünnschliff). Deutlich zu sehen ist das nach oben sternförmig zulaufende Exkretionsystem (Astrorhizenkanäle). |
|
